|
|
| Acta Bioquímica Clínica Latinoamericana |
| Control ponderal, leptina y neuropéptido "Y" |
|
El Comité de Redacción de Acta Bioquímica Clínica Latinoamericana ha seleccionado este trabajo publicado en Medical MAG vol 8 Nº 72, para su difusión a través de FABA-Informa
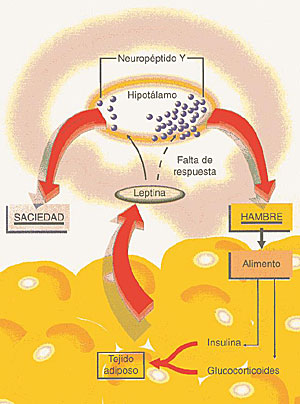
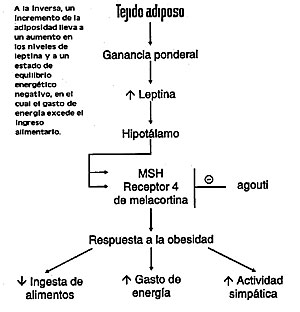
La capacidad de almacenar grandes cantidades de combustible de alta densidad energética, bajo la forma de tejido adiposo, permite a los vertebrados terrestres sobrevivir durante prolongados períodos de deprivación de alimentos. Con el fin de mantener estos depósitos energéticos sin experimentar alteraciones continuas en el tamaño y en la forma, el animal necesita alcanzar un equilibrio entre ingreso y gasto de energía. Una serie de informes de reciente publicación resaltan la importancia del neuropéptido Y, como también la de un receptor específico de melanocortina, como componentes clave del sistema que, en el seno de las estructuras cerebrales, regula el peso corporal. En la medida en que se ha incrementado la comprensión por parte de los científicos del diagrama de conexiones fisiológicas que permite el mantenimiento de un peso corporal sin grandes variaciones, las informaciones provenientes de este campo de investigación indican la existencia de diferentes vías de regulación de las respuestas biológicas que conducen al adelgazamiento y al aumento ponderal. Regulación cerebral Las nociones más elementales de la regulación cerebral de la ingestión alimentaria y el peso corporal datan de comienzos de este siglo, cuando se observó que, en algunos raros casos, el daño accidental de las estructuras cerebrales hipotalámicas tiene como resultado la obesidad. El compromiso de núcleos talámicos individuales en la regulación ponderal fue establecido en la década del 40 por Hetherington y Ranson. Estos autores, considerados entre los mejores neuroanatomistas de este siglo, realizaron experiencias en roedores a los cuales volvieron obesos produciéndoles lesiones en algunos núcleos seleccionados al azar dentro de la mencionada región cerebral. Esos datos neuroanatómicos, en combinación con estudios que documentaron la constancia del peso corporal en el tiempo indicaron que el equilibrio energético está controlado por centros integradores hipotalámicos. En la actualidad, comienzan a emerger los sustratos moleculares de ese sistema regulador hipotalámico. Un grupo de investigadores universitarios de Oregon y Arizona (EE.UU.) conducidos por Wei Fan, del Instituto de Investigación Biomédica Avanzada Vollum, demostraron que el receptor 4 de la melanocortina y su péptido ligando, la hormona estimulante de los melanocitos (MSH), son importantes en la patogenia de la obesidad en ratones que experimentaron una mutación de color y conocidos como "amarillo agouti" (AY). Por su parte, Erickson y col., del Instituto Médico Howard Hughes y del Departamento de Bioquímica de la Universidad de Washington (Seattle, EE.UU.), comunicaron mutaciones en el gen que codifica el neuropéptido Y cerebral, las que conducen a una reducción de la obesidad y de otras anormalidades que se observan en los ratones con mutaciones en el gen ob. Además de confirmar que la MSH y el neuropéptido Y se encuentran involucrados en la regulación del contenido de grasa corporal, estas novedades pueden tener gran importancia para dilucidar la estructura del sistema que regula el peso corporal. Ésta puede ser más sencilla de comprender si se consideran las respuestas fisiológicas a la pérdida de peso como funcionalmente diferentes de las respuestas al aumento de peso. Para desarrollar esta idea más extensamente es necesario formular algunas consideraciones respecto de la leptina. Gen o/b El concepto de que las anomalías genéticas contribuyen a la obesidad ha ido ganando mayor impulso con la identificación del gen ob y de su producto proteico, hacia 1994. La proteína ob, denominada leptina (del griego leptos, que significa delgado), se produce en el tejido adiposo y se piensa que actúa como una señal aferente de saciedad en un mecanismo de retroalimentación que afecta putativamente los centros del apetito y de la saciedad cerebrales. El efecto último de esa retroalimentación es la regulación de la masa de grasa corporal. En los ratones ob/ob, marcadamente hiperfágicos y obesos, el gen ob es mutante y no produce leptina; cuando se la administra en forma exógena, los animales paran de comer y pierden peso. Al parecer, esto no es lo que se produce en la mayor parte de los seres humanos obesos. Según Considine y otros investigadores de la Universidad Thomas Jefferson, de Filadelfia (EE.UU.), las concentraciones de leptina sérica y el nivel de RNA mensajero en los adipocitos de seres humanos obesos se encuentran elevados, y existe una correlación fuertemente positiva entre la concentración sérica de leptina y el porcentaje de grasa corporal, el índice de masa corporal y las concentraciones basales de insulina sérica. Estos resultados sugieren que los adipositos de los seres humanos producen leptina cuando se encuentra aumentada su masa y existe una resistencia a la acción de la leptina, de tal manera que se mantiene el aumento de la masa tisular adiposa (Fig. 1).
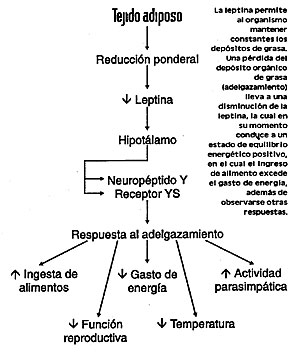
El problema en los sujetos obesos, por lo tanto, consiste en la disminución de la sensibilidad a la leptina, pero la naturaleza y las acciones del sistema efector para esta proteína se desconocen. Regulación de la secreción de leptina La leptina puede considerarse como la hormona de los adipocitos de la cual, excepto su relación con la masa grasa corporal, poco es lo que se sabe respecto a la regulación de su secreción. En seres humanos -según Considine y col.- y en animales, la restricción calórica reduce las concentraciones séricas de leptina y los niveles de RNA mensajero ob en el tejido adiposo, mientras que la realimentación aumenta esos niveles. El efecto del ayuno puede simularse mediante la administración de noradrenalina y el efecto de la alimentación mediante insulina o gluco-corticoides. La disminución en la expresión del gen ob en animales hambrientos que habían sido sometidos a ayuno, y su aumento por la administración de insulina y glucocorticoides (cuya concentración, al igual que la de insulina, aumenta después de las comidas) son compatibles con el concepto de que la leptina podría constituir un factor de saciedad. Puede avizorarse un sistema de retroalimentación en el cual la ingestión de alimentos podría desencadenar la secreción de insulina y glucocorticoides, por medio de la cual se favorecería la acumulación de grasa y luego la secreción de leptina, con la subsiguiente sensación de saciedad. Si la leptina es una hormona de saciedad y la ingestión de alimento es controlada por el cerebro (en particular por el hipotálamo) ¿cómo se relaciona la leptina con la ingestión de alimento? Una forma podría estar mediada por el péptido hipotalámico orexígeno, el neuropéptido Y (Fig. 2) (Fig.3).
Las elevadas concentraciones hipotalámicas del neuropéptido Y inducen la ingestión alimentaria, mientras que las bajas concentraciones tienen un efecto opuesto. En muchos modelos animales de obesidad, las concentraciones hipotalámicas de neuropéptido Y son elevadas y sus infusiones intraventriculares en ratas normales causan obesidad. El principal cambio metabólico que induce es el aumento de la secreción de insulina y glucocorticoides, los cuales llevan en su momento a la secreción de grasa, a la obesidad y a la resistencia a la insulina en el músculo, escalones que conducen a la diabetes mellitus no insulino dependiente. Las infusiones intraventriculares también causan un aumento de los niveles de RNA mensajero para leptina del tejido adiposo. Así, las concentraciones elevadas de neuropéptido Y en el cerebro producen la mayoría de las características de los síndromes de obesidad. La razón de estas elevadas concentraciones hipotalámicas de neuropéptido Y en roedores genéticamente obesos no está clara y sin duda difiere entre los diferentes síndromes de obesidad. En algunas cepas de ratas y ratones obesos, en los cuales las concentraciones de leptina sérica son elevadas, la acción de la leptina podría estar deteriorada, de tal manera que la inhibición de los factores orexígenos, el neuropéptido Y en particular, se deteriora, y las concentraciones hipotalámicas de éste son altas. En ratones ob/ob, dado que no se produce leptina, las concentraciones hipotalámicas de neuropéptido Y y de su RNA mensajero están elevadas. Cuando se administra leptina durante períodos prolongados a ratones ob/ob, se reduce su elevada ingestión de comida en igual medida que su peso corporal (como se notó más tarde), mientras que su nivel metabólico se eleva, como lo indica su consumo de oxígeno. Sin embargo, sus elevadas concentraciones de insulina, corticosterona y glucosa séricos, así como su actividad locomotriz, retornan al nivel normal. Los efectos beneficiosos de la leptina en esos ratones se acompañan con disminuciones marcadas en las concentraciones de neuropéptido Y. Neuropéptido Y: efector de la acción de la leptina De acuerdo con todo lo anterior, el neuropéptido hipotalámico Y podría tener un papel crucial como efector de la acción de la leptina. Que esta última actúa dentro del sistema nervioso central queda sugerido por la observación de que la dosis requerida para disminuir la ingestión alimentaria es menor cuando se administra por vía intraventricular que por la vía periférica. Hay coincidencia entre esta observación y la presencia en el tejido hipotalámico de sitios de unión de alta afinidad para la leptina. Por lo tanto, como sugieren los resultados de la experiencia de Considine y col. respecto de los sujetos obesos y por los resultados de los estudios realizados en varios modelos animales, un mecanismo fundamental de la obesidad es la insensibilidad a la acción de la leptina, presumiblemente en el hipotálamo. Se desconoce si ella se debe a mutaciones del gen para los receptores de la leptina en el cerebro, a anormalidades posreceptor en la transducción de la señal de la leptina o a otras anomalías en la función hipotalámica. Pese a estas incertidumbres, el hallazgo de esta hormona constituye un jalón de importancia en la comprensión de la fisiopatología de la obesidad. Para quienes han propuesto que ésta es el resultado de la desregulación de un sistema que liga el cerebro con la ingestión alimentaria y la masa de tejido adiposo, la leptina viene a llenar un vacío prolongado en ese sistema. También otros factores contribuyen a la disfunción de este sistema, por lo tanto la obesidad puede incluir hiperfagia inducida por neuropéptido Y, deficiencia en la producción o en la acción de neuropéptidos hipotalámicos anorexígenos e incremento de la secreción de insulina y glucocorticoides. Debido a que estos dos últimos estimulan la secreción de leptina, esta hormona representa una vía aferente que permite disminuir la actividad neuropeptidérgica Y y la reducción de la ingestión alimentaria. Así, la masa normal de grasa corporal puede mantenerse por medio de un sistema de retroalimentación central-periférico que en los individuos obesos se encuentra alterado. En ausencia de leptina (como en los ratones ob) los animales no pueden restringir la ingestión alimentaria y se tornan obesos. Estos animales padecen un síndrome complejo, en el cual se observa la mayoría de los síntomas de inanición, o sea función reproductiva anormal, hipotermia, detención del crecimiento, anomalías hormonales y disminución de la actividad. Por consiguiente, los animales ob parecen existir en un estado de percepción de inanición y, en consecuencia, se tornan obesos. Si se los trata con leptina, se corrige la obesidad y las anormalidades que indican la respuesta a la inanición se revierten. Por otra parte, si se tratan animales normales con leptina, pueden prevenirse algunos de los síntomas resultantes de la limitación de la ingestión de alimentos. El estudio de Erickson y col. considera la producción de NPY como la resultante de un bajo nivel de leptina en el marco de esta respuesta biológica. Se pensó que el gen NPY estaba involucrado en el desarrollo del fenotipo ob, debido a que su inyección en el cerebro aumenta la ingestión de alimentos y el peso corporal. Mas aún, los niveles endógenos de mRNA de NPY son mucho más elevados en ratones ob que en normales, pero disminuyen después del tratamiento con leptina. En forma sorpresiva, los roedores con mutaciones en el gen NPY tienen peso normal y responden también normalmente a la leptina exógena. Pero cuando Erickson y col. cruzaron ratones con NPY descendido y ratones ob encontraron que de cualquier modo el NPY se encontraba involucrado en la regulación del peso corporal. |
|
|